

Добавить статью
| Главная » Статьи » Прудовое рыбоводство. » Промышленное производство |
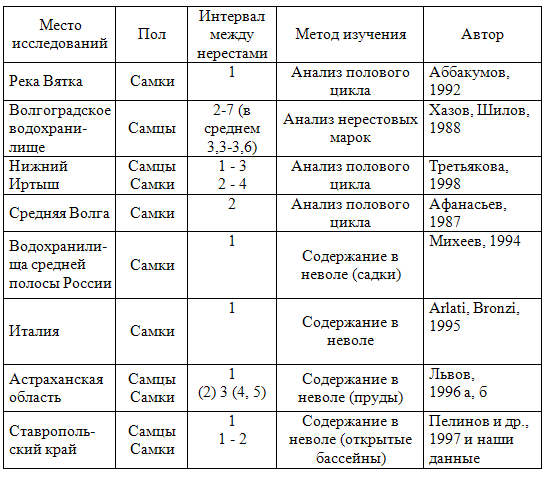
Научно-технический бюллетень лаборатории ихтиологии ИНЭНКО. 1999. № 2. С.20-38. УДК 597.442 Обобщению литературных данных о цикличности размножения осетровых были посвящены обзоры З.С.Коробочкиной (1964) и наш (Подушка, 1989). С момента публикации последнего обзора прошло уже 10 лет, и в литературе появилось довольно много новых сведений, требующих своевременного обобщения и анализа. Настоящая работа является дополнением к предыдущему обзору (Подушка, 1989), в ней рассмотрены преимущественно материалы, опубликованные в последние годы. Основными методами определения длительности межнерестовых интервалов у осетровых являются: выявление нерестовых марок на спилах лучей грудного плавника, изучение половых циклов, мечение рыб в естественных популяциях и наблюдение за производителями, содержащимися в искусственных условиях. В предыдущем обзоре (Подушка, 1989) подробно рассмотрены преимущества и недостатки каждого из названных методов. Наиболее распространенным методом изучения периодичности размножения осетровых в естественных условиях является выявление нерестовых марок на спилах плавниковых лучей. Основополагающей работой в этом направлении считается исследование Г.Руссова (Roussow, 1957), который предположил, что созревание половых продуктов у осетровых сопровождается замедлением темпа роста. В результате этого, по мнению автора, на спилах костных лучей грудных плавников у половозрелых рыб периодически появляются пояса сближенных годовых слоев, чередующиеся с сериями широких приростов. Анализируя эти структуры, Г.Руссов определял у рыб количество нерестов и длительность межнерестовых интервалов. Впоследствии эту методику использовали многие ихтиологи при изучении биологии различных видов осетровых. Недавно группа канадских исследователей (Guénette et al., 1992) предприняла попытку провести ревизию работы Г.Руссова. Объектом исследования был озерный осетр Acipenser fulvescens — тот же вид, с которым работал Г.Руссов. Авторы изучили спилы лучей у 125 самок этого вида возрастом 24 года и старше, используя несколько приемов для анализа приростов годовых колец. У большинства рыб не прослеживалось какой-либо связи между степенью зрелости половых желез в момент вылова и шириной приростов последних пяти лет на спилах лучей грудных плавников. Авторы пришли к заключению, что методика выявления и анализа нерестовых марок малополезна при изучении периодичности размножения озерного осетра в бассейне реки Св.Лаврентия. Особо следует остановиться на вопросе о периодичности созревания русского осетра Acipenser gueldenstaedti Азовского моря. Анализ созревания этого вида, сделанный Э.В.Макаровым (1970) по нерестовым маркам на спилах лучей грудного плавника, показал, что единично первые самки осетра созревают в восьмилетнем возрасте, основная масса самок созревает в 13-17 лет, а межнерестовые интервалы у рыб обоих полов длятся от трех до девяти лет, составляя для большинства самок между первым и вторым икрометаниями 5-6 лет, между вторым и третьим — 4-5 лет и между третьим и четвертым — 3-4 года. Впоследствии Ю.И.Реков (1983), анализируя темп созревания азовского осетра по нерестовым маркам за более поздние годы, отметил аномально низкий возраст первого созревания у ряда особей (с шести лет у самок), что не соответствовало реальному возрастному составу нерестового стада этого вида. Сходная картина была отмечена и для севрюги Acipenser stellatus. Автор предположил, что часть структур на спилах лучей плавников, принимаемых за нерестовые марки, в действительности таковыми не являются. Позже вывод о том, что первая нерестовая марка у части особей азовских осетровых является ложной, был подтвержден на основании изучения полового цикла азовского осетра (Корниенко и др., 1988). Вероятно, у части неполовозрелых осетров и севрюг в Азовском море по каким-то причинам происходило временное замедление роста, проявляющееся на спилах лучей грудных плавников в виде сближенных годовых слоев, напоминающих нерестовые марки. В этой связи интересно упомянуть данные из работы Ю.В.Алтуфьева и Л.Ф.Львова (1972). Эти авторы отметили, что при выращивании в прудовых условиях у осетра, севрюги и белуги в возрасте 4-5 лет (то есть примерно в возрасте появления ложных нерестовых марок у азовских осетровых) наступает "критический период”, когда наблюдается резкий спад весовых и линейных приростов, что вызывает сужение промежутков между годовыми кольцами на спилах лучей грудного плавника. Г.Ф.Металлов (1994) указывает на уменьшение ширины годовых колец у каспийских осетровых в периоды острых экологических ситуаций в море (осолонение и загрязнение). В публикациях 1988-1989 гг. сотрудники АзНИИРХа (Корниенко и др., 1988, 1989) определяли межнерестовые интервалы для азовского осетра в 4-5 лет для самцов и 5-6 лет для самок. В более поздней статье (Корниенко и др., 1996 а) даются более низкие оценки — 3-5 лет для обоих полов. Для азовской севрюги Г.Г.Корниенко с соавторами (1996 а) указывают длительность межнерестовых интервалов в 2-4 года для обоих полов, что также ниже прежних оценок Э.В.Макарова (1970) и С.В.Тихоновой (1972). Повторные нересты у каспийской белуги Huso huso, по оценкам В.М.Распопова (1992, 1993), основанным на анализе линейного и весового роста, наблюдаются у самок с интервалом в 4-8 лет, у самцов — в 4-7 лет. При благоприятных условиях белуга, по мнению этого автора, может заходить в Волгу на нерест до 9 раз. Дж.Кемпинджер (Kempinger, 1988), ссылаясь на неопубликованные данные Маньена и Прежеля (Magnin, Priegel), указывает межнерестовые интервалы для озерного осетра Acipenser fulvescens в естественных водоемах 4-6 лет для самок и 2 года для самцов. В последние годы сведения о периодичности размножения осетровых пополнились данными, полученными в рыбоводных хозяйствах, поскольку масштабы культивирования этих рыб существенно увеличились. Это касается прежде всего сибирского (ленского) осетра Acipenser baeri, стерляди A. ruthenus, адриатического осетра A. naccarii и белого осетра A. transmontanus. В табл.1 приведены опубликованные за последние годы сведения о длительности межнерестовых интервалов у стерляди. При указании периодичности размножения цифра 1 в этой и следующей таблице означает ежегодное созревание, цифра 2 — созревание с двухлетним интервалом, 3 — с трехлетним и так далее. Данные таблицы показывают, что оценки периодичности размножения стерляди в естественных водоемах, даваемые разными авторами, существенно расходятся, а в условиях культивирования как самцы, так и самки могут созревать ежегодно. Л.Ф.Львов (1996 а) приводит данные о влиянии условий содержания и кормления прудового маточного стада стерляди на межнерестовые интервалы производителей. При высоких плотностях посадки (450-750 экз./га) и содержании только на естественной кормовой базе даже у самцов созревают ежегодно не более 30% особей. При меньших плотностях посадки и дополнительном кормлении ежегодно созревают более 90% самцов. Длительность повторных созреваний самок стерляди в таких условиях колеблется от двух до пяти лет (наиболее часто межнерестовый интервал составляет три года). Таблица 1 Длительность межнерестовых интервалов у стерляди
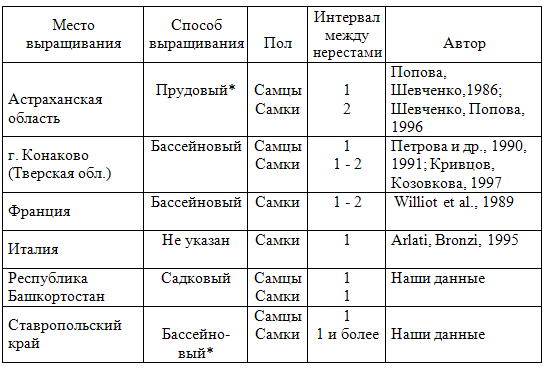
В табл.2 представлены данные о периодичности размножения сибирского (ленского) осетра. Самцы этого вида созревают ежегодно, как при содержании в прудах южных районов России, так и в тепловодных бассейновых и садковых хозяйствах, самки же в прудах имеют преимущественно двухлетний половой цикл, а в бассейновых и садковых хозяйствах большая или меньшая часть рыб может созревать ежегодно. В естественных условиях межнерестовые интервалы у производителей этого вида составляют не менее 2-3 лет (Кошелев и др., 1988, 1989). Перерывы между последовательными нерестами у самок адриатического осетра в условиях культивирования длятся от одного до трех лет (Белов и др., 1990; Bronzi et al., 1994; Arlati, Bronzi, 1995). В сформированном в искусственных условиях в США маточном стаде белого осетра большинство самцов имеют годичный половой цикл, а большинство самок — двухлетний (Doroshov et al., 1997). Отловленные в естественных водоемах самки русского осетра Acipenser gueldenstaedti, помещенные после прижизненного получения от них икры в пруды, повторно созревают через 3-4 года (Попова и др., 1997). Самцы этого вида, выращенные до половозрелости в индустриальных рыбоводных хозяйствах, по нашим наблюдениям, созревают обычно ежегодно. Таблица 2 Длительность межнерестовых интервалов у сибирского осетра в рыбоводных хозяйствах (по оценкам разных авторов)
Примечание* — естественный температурный режим. В остальных случаях — выращивание с использованием подогретой воды. Мойази Амири с соавторами (Mojazi Amiri et al., 1996 a, b) изучали половой цикл у одиннадцатилетних производителей бестера (гибрид белуги со стерлядью) в двух бассейновых рыбоводных хозяйствах Японии, различающихся температурным режимом. У рыб обоих полов в течение года регулярно брали биопсийные пробы половых желез для гистологического анализа. Исследование показало годичный половой цикл у самцов в обоих хозяйствах. У самок бестера отдельные особи существенно различались между собой по скорости оогенеза. Авторы работы пришли к выводу, что для точного заключения о длительности межнерестовых интервалов у самок период наблюдений должен быть не менее трех лет. Однако приведенные в статье материалы позволяют предположить, что минимальный промежуток между последовательными нерестами у самок бестера может быть два года. У веслоноса Polyodon spathula, представителя родственного осетровым семейства Polyodontidae, при выращивании его в водоемах юга России созревание самцов происходит ежегодно, а самок — раз в два года (Виноградов и др., 1996; Мельченков и др., 1996; Илясова, 1997). По данным В.В.Архангельского с соавторами (1997), самок веслоноса для получения икры можно использовать не менее пяти раз. Довольно противоречивые сведения имеются о зависимости между длительностью межнерестовых интервалов и приростами массы тела производителей. По данным И.А.Бурцева с соавторами (1981), у самок стерляди, созревающих ежегодно, наблюдалось значительно меньшее увеличение массы тела за год, чем у рыб, имеющих более длительные промежутки между нерестами, то есть имела место обратная зависимость между генеративным и соматическим ростом. Однако в более ранней работе этот же автор (Бурцев, 1967) показал, что перевод гибридов белуги со стерлядью с полуголодного существования к интенсивному откорму приводит одновременно к резкому усилению весового и линейного роста и активизации гаметогенеза. В работе Л.Ф.Львова (1996 а) отмечена аналогичная картина для стерляди. При неблагоприятных условиях содержания невысокие приросты массы тела и низкая упитанность коррелировали с невысоким процентом ежегодно созревающих самцов. Улучшение условий содержания приводило к увеличению упитанности и приростов рыб и сопровождалось ежегодным созреванием большинства самцов. Таким образом, в данном случае имела место прямая связь между генеративным и соматическим ростом. Сходную картину мы наблюдали у самок ленского осетра в рыбоводном цехе Алексинского химического комбината. Среди особей, от которых весной 1996 г. была получена икра, часть рыб повторно созрела к весне 1997 г. Оказалось, что у этих самок наблюдалось значительное увеличение массы тела, тогда как у пропускающих нерестовый сезон рыб приросты были очень малы. По-видимому, соотношение между генеративным и соматическим ростом у осетровых является довольно изменчивым признаком, который сильно зависит от многих факторов. В связи с этим есть серьезные основания полагать, что и чередование сближенных и широких годовых колец на спилах лучей грудного плавника (нерестовые марки), отражающее соматический рост, с генеративным обменом может быть связано самым разнообразным образом. В последние годы появилась серия публикаций, посвященных выявлению и описанию различных аномалий гаметогенеза, которые потенциально могут сказываться на сроках достижения рыбами половой зрелости, длительности повторных созреваний и воспроизводительной способности. В большинстве случаев, обнаруживая какие-либо отклонения от нормы, авторы объясняют их появление загрязнением окружающей среды или другими последствиями человеческой деятельности. Среди широкого спектра описанных аномалий гаметогенеза в наибольшей степени оказывать влияние на периодичность размножения осетровых, по-видимому, могут лишь случаи массовой гибели половых клеток. Впервые дегенерация превителлогенных ооцитов как массовое явление была описана В.Г.Свирским (1958) у амурского осетра Acipenser schrenki. Впоследствии этот же автор отметил тотальную дегенерацию и у калуги Huso dauricus (Свирский, 1979). Дегенеративные изменения в яичниках, по данным В.Г.Свирского, завершаются интенсивной пролиферацией клеток герминативного эпителия и митотической активностью оогониев. Дегенеративные и восстановительные процессы в яичниках этот автор рассматривает как своеобразную видовую адаптацию, обеспечивающую сохранение репродуктивных функций особей, оказавшихся по тем или иным причинам в стрессовых ситуациях. Массовую гибель ооцитов перед переходом к периоду протоплазматического роста обнаружил И.А.Бурцев (1962) у многих самок гибрида русского осетра со стерлядью. Гибель ооцитов не нарушала процесса размножения оогониев, и в яичниках всегда присутствовали половые клетки разных фаз развития — от оогониев до ооцитов периода синаптенного пути и единичных ооцитов периода протоплазматического роста. Нарушения оогенеза И.А.Бурцев объясняет стерильностью данного гибрида, хотя среди исследованных им рыб были особи и с нормальным состоянием яичников. В связи с тем, что позже массовая дегенерация ооцитов ранних фаз развития была отмечена как весьма распространенное явление не только у амурских осетровых и гибридов, но и у сибирского осетра в различных частях ареала (Акимова, Рубан, 1991, 1992; Рубан, Акимова, 1991, 1993; Акимова и др., 1995; Akimova, Ruban, 1995), можно предполагать, что она не представляет собой патологического процесса, а является характерной особенностью функционирования половых желез осетровых в суровых климатических условиях. Встречается это явление и у азовских (Корниенко и др., 1994, 1996 б; Моисеева и др., 1997) и каспийских осетровых (Романов и др., 1990; Саенко, 1998). И.И.Саенко (1998) связывает повреждения половых клеток резервного фонда с возрастным угасанием репродуктивной функции у русского осетра. У некоторых видов костистых рыб массовая дегенерация превителлогенных ооцитов может быть нормой полового цикла (Свирский, 1990; Сакун, Свирский, 1992). | |
| Просмотров: 1632 | Рейтинг: 0.0/0 |
| Всего комментариев: 0 | |
